
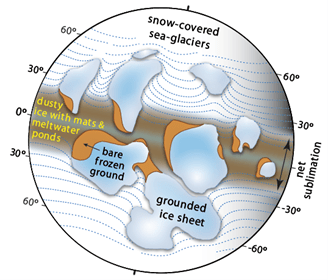



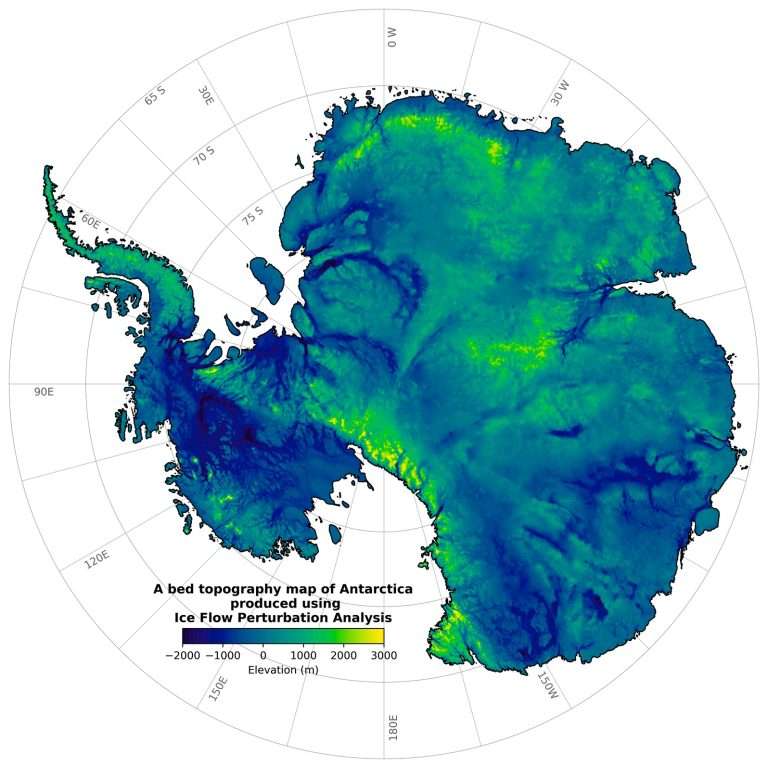
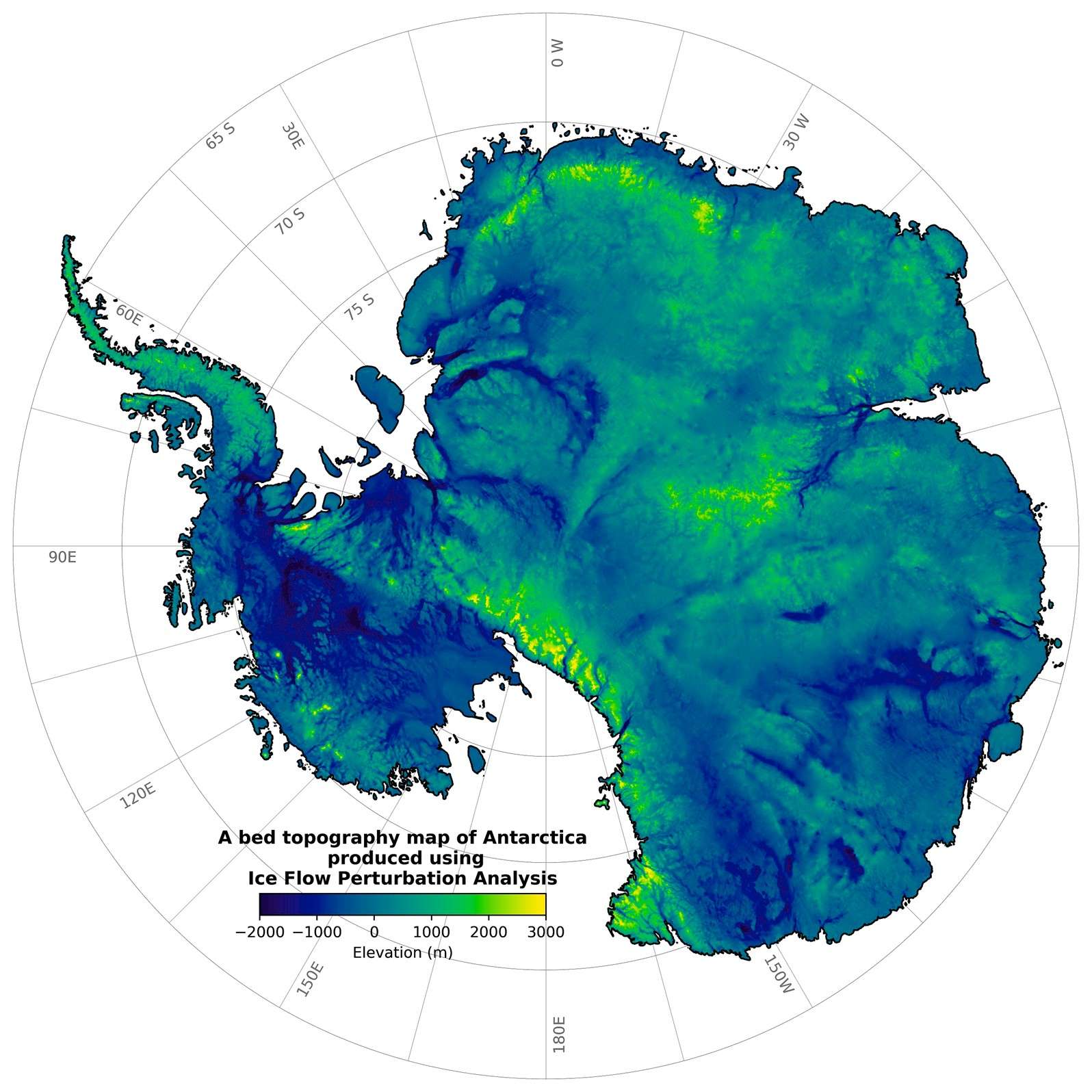

A native of Toronto, Hoffman was a Senior Research Scientist with the Geological Survey of Canada (1969-92), Sturgis Hooper Professor of Geology at Harvard University (1994-2008), and Professor of Geology at The University of Victoria (1992-94) where he has been an Adjunct Professor since 2008. He was an early proponent of Precambrian plate tectonics, based on field research in northern Canada (1961-91), and the leading advocate of Snowball Earth, based on field research in northern Namibia (1993-2023). He is a recipient of career-achievement awards from learned societies on four continents, most recently the 2024 Kyoto Prize for Basic Sciences from the Inamori Foundation of Japan.

Can Egypt lower emissions and energy costs at the same time? This study shows how...

Can System Dynamics modelling help farmers and policymakers avoid systemic water crises? This research investigates...

Can urban growth make heatwaves deadlier for millions? A Europe-wide study uncovers how cities amplify...

Did simple fatty acid membranes drive early evolution?

Could future cyclones be even deadlier? Researchers at IIT Kharagpur demonstrate how warmer oceans and...